Frontiere în genetică
Introducere
Multe trăsături sunt reglementate și coordonate de mai multe gene și condiții de mediu. În natură, astfel de trăsături evoluează continuu sub presiunea selecției locale și a proceselor neutre, conducând la o diversitate bogată de varietăți fenotipice și de strategii de adaptare la mediu. Deja Dobzhansky (1964) a descris bogata diversitate din lumea vie ca fiind rezultatul diversității genetice, al eterogenității mediului și al adaptărilor care evoluează la interacțiunea dintre biologia moleculară și cea organică. Mecanismele moleculare în evoluție au fost deosebit de dificil de caracterizat. În primul rând, trebuie să facem legătura între genotip și fenotip. Această legătură constă în rețele extinse și complexe de interacțiune genetică (Ayroles et al., 2009; Lehner, 2013). În al doilea rând, trebuie să identificăm modificările moleculare responsabile de adaptările fenotipice. Acest lucru depinde de elucidarea variației genetice și a modificărilor genetice care pot apărea oriunde în rețelele de interacțiune genetică (de exemplu, Edwards et al., 2009).
Manifestarea variației genetice în rețelele de gene este foarte complexă. Modificările la nivelul unei singure gene pot afecta activitatea și chiar topologia întregii rețele genetice (de exemplu, Knight et al., 2006). Genele și rețelele de interacțiune genetică sunt adesea pleiotropice și reglează diverse trăsături și procese, ceea ce implică faptul că modificările într-o singură genă se pot manifesta în mai multe trăsături (Stearns, 2010). Invers, variațiile alelice în mai multe gene pot contribui la variația unei anumite trăsături fenotipice (Manolio et al., 2009). În plus, epistazia este omniprezentă, ceea ce implică faptul că variațiile alelice la mai mulți loci pot afecta efectele fenotipice ale fiecăruia dintre ele (Phillips, 2008; Mackay și Moore, 2014; Moore și Williams, 2015). Aceste considerații privind baza genetică a evoluției nu sunt noi și sunt studiate de câteva decenii (de exemplu, Wagner și Altenberg, 1996). Genetica directă și cea inversă au avut un mare succes în elucidarea funcțiilor unor gene unice sau mutații pentru o anumită trăsătură (Nagy et al., 2003). Cu toate acestea, aceste tehnici sunt limitative atunci când se studiază complexitatea rețelelor de interacțiune moleculară care stau la baza unui fenotip sau mecanismele moleculare ale evoluției trăsăturilor complexe.
Dezvoltarea tehnologiei genomice a reprezentat un impuls major pentru capacitatea noastră de a studia complexitatea genetică a trăsăturilor fenotipice și evoluția acestora (Stapley et al., 2010). Combinarea acestor tehnici cu abordările clasice ale geneticii ne permite să evaluăm funcționalitatea variației genetice pentru trăsăturile fenotipice (Storz și Wheat, 2010). Primele studii genomice pe mai multe organisme model au evidențiat faptul că adaptările evolutive, chiar și pentru condiții de mediu specifice, guvernează în general multe gene sau loci, precum și reglarea dinamică a modelelor de expresie genetică (Gasch et al., 2000; Fay et al., 2004; Pedra et al., 2004). În deceniul următor, numeroase studii au utilizat genomica pentru a identifica genele și proteinele care contribuiau la anumite trăsături și interacțiuni ecologice. Inițial, costurile și timpul necesar pentru secvențierea unui singur genom erau încă foarte restrictive. Cu toate acestea, secvențierea de generație următoare a făcut posibilă secvențierea genomurilor a mult mai multor specii și a mult mai multor indivizi per specie. Aceasta este o resursă formidabilă pentru studierea evoluției, deoarece ne permite, pentru prima dată, să cartografiem schimbările la nivelul întregului genom în timpul evoluției.
Tehnologia genomică are un potențial uriaș pentru a ne îmbunătăți cunoștințele despre procesele evolutive. Abordările comparative au fost aplicate pentru a cartografia modificările în secvențele genomului sau în rețelele de interacțiune genetică la scări temporale evolutive lungi (Drosophila 12 Genomes Consortium et al., 2007; Nowick et al., 2009; Jones et al., 2012). Abordările de selecție experimentală sau de evoluție experimentală, urmate fie de secvențierea transcriptomică, fie de secvențierea genomului, au fost utilizate pentru a cartografia schimbările evolutive la scări temporale mult mai scurte (Hunt et al., 2010; Turner et al., 2011; Wertheim et al., 2011; Tenaillon et al., 2012; Linnen et al., 2013; Jalvingh et al., 2014). Aceste studii au arătat, de exemplu, modul în care duplicările de gene, mutațiile și divergențele puternice de secvențe într-un subset mic de gene pot avea un impact profund asupra activității transcripționale a unor rețele mari de interacțiune genetică și a mai multor trăsături fenotipice.
În această perspectivă, prezint descoperirile și evoluțiile recente privind bazele genomice ale evoluției, folosind imunitatea în evoluție ca studiu de caz. Imunitatea este o trăsătură care evoluează rapid, ceea ce o face potrivită pentru a studia baza genomică a proceselor evolutive (Obbard et al., 2009; Sironi et al., 2015). În plus, rețelele moleculare din imunitate au fost destul de bine caracterizate datorită importanței lor pentru sănătatea umană (Schadt, 2009; Lazzaro și Schneider, 2014; Zak et al., 2014). În primul rând, voi rezuma foarte pe scurt înțelegerea noastră actuală cu privire la rețelele moleculare ale răspunsurilor imune înnăscute și la procesele de selecție care acționează asupra răspunsurilor imune. Apoi, descriu modificările genomice, asociate cu câștigarea, pierderea și modularea anumitor aspecte ale răspunsurilor imune la Drosophila. În cele din urmă, propun direcții viitoare pentru a studia arhitectura genetică a trăsăturilor complexe și a proceselor evolutive.
Răspunsuri imune
Sistemul imunitar constă într-o combinație de procese fiziologice care acționează împreună în apărarea împotriva agenților patogeni și a paraziților. Imunitatea înnăscută este o trăsătură străveche care poate fi întâlnită la toate organismele pluricelulare, în timp ce vertebratele posedă, de asemenea, imunitate dobândită. Ambele sisteme imunitare combină componente celulare și umorale: componenta celulară cuprinde celule specializate care asigură o funcție de protecție. Aceasta include, clase de celule sanguine pentru fagocitoza microbilor, încapsularea corpurilor străine mai mari sau recunoașterea antigenelor (în cazul imunității dobândite) și mucoasa intestinală cu celule epiteliale care formează o barieră fizică și pot secreta compuși defensivi. Componenta umorală constă în eliberarea de factori extracelulari care combat agenții patogeni invadatori, adesea din țesuturi sau populații celulare specializate. Aceasta include eliberarea de peptide antimicrobiene (AMP) de către ficat (sau corpul adipos la nevertebrate) și celulele epiteliale intestinale, specii reactive de oxigen în celulele fagocitare și epiteliale și anticorpi din celulele albe din sânge (în imunitatea dobândită; Lemaitre și Hoffmann, 2007; Buchmann, 2014).
Rețele complexe de interacțiuni moleculare coordonează răspunsurile imune (Figura 1A). Aceleași căi sunt centrale pentru răspunsurile imune de la nevertebrate la vertebrate, ceea ce implică o conservare puternică a elementelor de bază ale rețelelor moleculare în imunitate (Silverman și Maniatis, 2001; Evans et al., 2003; Buchmann, 2014). O varietate de molecule receptoare pot recunoaște agenții patogeni sau paraziții, de exemplu, pe baza modelelor moleculare asociate agenților patogeni (de exemplu, lipopolizaharidele specifice membranelor bacteriene). Odată ce receptorii sunt activați, aceștia induc căi specifice de transducție a semnalelor, cum ar fi căile Toll, Imd și Jak/Stat (analizate în Lemaitre și Hoffmann, 2007; Buchmann, 2014). Aceste căi constau din proteaze, kinaze, citokine și alte proteine care, în cele din urmă, activează factorii de transcripție și cofactori. Inducerea acestor factori de transcripție duce la producerea de molecule efectoare umorale (de exemplu, AMP) și poate induce proliferarea și diferențierea celulelor implicate în imunitate. Producția de diferite clase de celule sanguine este un aspect proeminent al componentei celulare a răspunsului imunitar, atât în imunitatea înnăscută, cât și în cea dobândită. În aceste celule sanguine, cascadele de transducție a semnalelor sunt, de asemenea, reglementate pentru a induce proprietăți celulare și proteine care realizează eliminarea parazitului. Pentru a regla intensitatea, specificitatea, momentul și durata răspunsurilor imune, rețelele moleculare sunt modulate de citokine, proteaze și de interacțiunea încrucișată cu alte căi de semnalizare (Liew et al., 2005; Aggarwal și Silverman, 2008). Aceasta include, de asemenea, diverse rețele de reglementare post-transcripțională (Ivanov și Anderson, 2013; Carpenter et al., 2014).
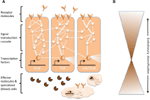
Figura 1. Reprezentarea schematică a rețelelor genetice în imunitate. (A) Mai multe rețele interconectate coordonează răspunsurile la o provocare imunitară. Aceste rețele sunt formate din proteine (reprezentate prin cercuri) care interacționează între ele într-o cascadă de transducție a semnalelor pentru a regla expresia factorilor de transcripție (reprezentați prin hexagoane). Activarea căilor de transducție a semnalelor de bază (de exemplu, IMD, Toll sau Jak/Stat, indicate prin linii groase între proteine) duce la producerea de molecule efectoare, cum ar fi peptidele antimicrobiene (reprezentate prin simboluri în formă de plăcintă) și la proliferarea și diferențierea celulelor specializate (sanguine) (figuri în formă de nor). Moleculele receptoare extracelulare și cele legate de membrană (figuri în formă de lună) induc aceste căi. Activitatea poate fi modulată în continuare de multe alte proteine care interacționează cu căile și intră în legătură încrucișată cu alte căi și rețele genetice (indicate prin linii subțiri între proteine). (B) Componentele centrale ale rețelelor genetice în imunitate, de exemplu, factorii de transcripție și proteinele în contact direct cu acești factori de transcripție, sunt adesea puternic conservate între phyla. Diversificarea evolutivă se regăsește mai mult spre periferiile rețelelor.
Propulsorii schimbărilor evolutive în răspunsurile imune sunt efectele combinate ale costurilor de fitness ridicate ale infecției asupra gazdelor, costurile imunității, diversitatea bogată a agenților patogeni și a paraziților care amenință gazdele și cursele dinamice de arme co-evolutive dintre gazde și agenți patogeni (Schmid-Hempel, 2003). Oricare sau toate aceste aspecte pot acționa în mediul local al gazdei și pot conduce la presiuni de selecție puternice. Cu toate acestea, tipul de selecție variază de la direcțională, la purificare sau echilibrare, în funcție de costurile și beneficiile pe care populația gazdă le experimentează în mediul său local. De exemplu, un agent patogen foarte virulent care invadează o comunitate locală poate provoca o selecție selectivă sau o selecție direcțională pentru anumite alele de rezistență, deoarece numai gazdele cu aceste alele pot contribui la generațiile următoare. Alternativ, o comunitate de agenți patogeni diversă sau în co-evoluție poate determina o selecție dependentă de frecvență sau o selecție de echilibrare, favorizând menținerea variației genetice. Evoluția răspunsurilor imune reflectă atât aceste dinamici co-evolutive cu parazitul, cât și costurile fiziologice și ecologice ale sistemului imunitar (Kraaijeveld et al., 2002; Rolff și Siva-Jothy, 2003; Schmid-Hempel, 2005; Lazzaro și Little, 2009).
Bazele genomice ale schimbărilor evolutive în imunitatea Drosophilei
În timp ce mecanismul central al răspunsurilor imune este puternic conservat, mai multe componente ale rețelelor moleculare extinse pot evolua rapid sau se pot diversifica (Figura 1B). La Drosophila, au fost raportate schimbări evolutive rapide pentru receptorii și efectorii răspunsului imunitar (Sackton et al., 2007; Obbard et al., 2009; Salazar-Jaramillo et al., 2014). Aceste molecule acționează la interfața dintre gazdă și agentul patogen și, prin urmare, sunt esențiale pentru recunoașterea de către gazdă a unui organism invadator și pentru a media direcționarea și efectele antagoniste ale răspunsului imunitar asupra agentului patogen. În același timp, parazitul este supus selecției pentru a trece neobservat, pentru a evita sau a atenua efectele antagoniste ale răspunsului imunitar. Prin urmare, se așteaptă o dinamică de tip Red Queen pentru aceste molecule de la interfață și pentru cele care modulează răspunsurile imune. Fiecare parte încearcă să câștige avantajul în cursa antagonistă a armelor, determinând reciproc modificări în rețelele genetice ale părților. Diversificarea moleculelor receptoare, modulatoare și efectoare se realizează în cea mai mare parte prin duplicări de gene și schimbări rapide de secvențe (Drosophila 12 Genomes Consortium et al., 2007; Sackton et al., 2007; Salazar-Jaramillo et al., 2014).
Am studiat răspunsul imunitar al Drosophilei împotriva viespilor parazitoide ca sistem model pentru a înțelege baza genomică a proceselor evolutive. Larvele de Drosophila sunt gazda unei varietăți de specii de parazitoizi care depun un ou în aceste larve (Fleury et al., 2009). Odată ce oul parasitoid eclozează (∼2-4 zile după atacul parasitoidului, în funcție de specia de parazitoid și de temperatură), larva parasitoidului începe să se hrănească cu gazda și o ucide. Unele specii de Drosophila dispun de un mecanism de apărare împotriva parazitoizilor printr-un răspuns imunitar înnăscut, numit încapsulare melanotică. Acest răspuns imun este format din componente celulare și umorale care acționează împreună pentru a sechestra și ucide oul parazitului. Atacul parazitului declanșează căi de transducție a semnalelor imune care induc (i) proliferarea și diferențierea a două clase de hemococite (adică celulele sanguine ale insectelor) care aderă la oul parazitului și între ele și (ii) depunerea de melanină pe oul parazitului și pe capsula celulară din jurul oului parazitului (Lemaitre și Hoffmann, 2007). Gazda trebuie să finalizeze încapsularea completă și melanizarea înainte ca oul de parazit să eclozeze pentru a supraviețui infestării cu parazit.
Înainte de era genomică, fuseseră identificate mai multe gene implicate în răspunsul imunitar împotriva viespilor parazitoide. Căile Toll și Jak/Stat fuseseră identificate ca fiind componente centrale ale proliferării și diferențierii hemocitelor, iar calea profenoloxidazei pentru melanizare (revizuite în Brennan și Anderson, 2004). Două studii de microarray au fost apoi efectuate pentru a identifica gene suplimentare potențial implicate în încapsularea melanotică după atacul parazitului. Această abordare a evidențiat multe gene suplimentare care nu fuseseră asociate anterior cu răspunsul imunitar împotriva parazitoidelor și a dezvăluit calendarul de acțiune al acestora (Wertheim et al., 2005; Schlenke et al., 2007). Studiile au scos la iveală mai multe grupuri coordonate și coerente din punct de vedere funcțional de gene care au fost temporar reglementate în sus sau în jos în timpul unei părți a răspunsului imunitar (Wertheim et al., 2005). În mod interesant, s-a demonstrat că mecanismele de virulență a două specii de parazitoizi diferă în modul în care au interferat în rețeaua genetică a răspunsurilor gazdelor: o specie a eliminat activarea inițială a întregii rețele, în timp ce o altă specie a vizat ultimul pas din cascadă (Schlenke et al., 2007).
Speciile de Drosophila diferă în mare măsură în ceea ce privește imunitatea împotriva parazitoizilor. Unele specii sunt complet sensibile la parazitoizi și acest lucru a fost raportat ca o deficiență imunitară (Eslin și Doury, 2006). Cu toate acestea, o examinare mai atentă a arătat că rezistența la parazitoizi nu este împărtășită în mod obișnuit de toate speciile de Drosophila, ci este limitată la câteva clade. Am arătat că, în una dintre aceste clade, subgrupul melanogaster, câștigul evolutiv al rezistenței la parazitoizi a fost asociat cu câștigarea unui nou tip de celule sanguine, lamelocitele, care este, de asemenea, restrâns la aceeași cladă (Salazar-Jaramillo et al., 2014). Unele specii de Drosophila din afara subgrupului melanogaster pot, de asemenea, să încapsuleze ouăle de parazitoizi, dar acestea par să fi evoluat diferite tipuri de celule sanguine pentru răspunsul de încapsulare (Havard et al., 2012; Márkus et al., 2015). Răspunsul imunitar împotriva parazitoizilor a evoluat independent la diferiți taxoni de insecte, adesea cu mecanisme și tipuri de celule sanguine ușor diferite (Lavine și Strand, 2002). Astfel, evoluția sistemului imunitar înnăscut include adăugarea de noi componente sau „module”. Acest lucru ridică întrebarea cum se modifică genomul în timpul dobândirii unui nou modul.
Genomica comparativă a arătat că, în ciuda obținerii unui nou tip de celule sanguine în subgrupul melanogaster, genele cunoscute pentru diferențierea lamelocitelor sunt în mare parte conservate în întreaga filogenie. De asemenea, speciile care nu produc lamelocite ca răspuns la atacul parazitoidelor posedă aceste gene care sunt necesare pentru diferențierea lamelocitelor. Mai mult, aceste gene prezintă puține divergențe sau semnături de selecție, în timp ce acest lucru ar fi de așteptat pentru genele care au obținut o funcție nouă. Acest lucru indică faptul că că căile de transducție a semnalului existente pentru diferențierea hemocitului sunt modulate de rețeaua de interacțiune genetică din jur pentru a produce un nou tip de celule sanguine în subgrupul melanogaster. Această cooptare a căii de bază existente de proliferare a hemocitului se realizează probabil prin adăugarea altor componente sau a unor componente noi în rețeaua de interacțiune genetică (Salazar-Jaramillo et al., 2014). Am identificat mai multe gene noi care au apărut în jurul momentului de achiziție a lamelocitelor și care sunt exprimate diferențiat în timpul răspunsului imunitar împotriva parazitoizilor, inclusiv molecule receptoare și proteaze de tip serină (Salazar-Jaramillo et al., 2014). Formulăm ipoteza că în special endopeptidazele de tip serină pot juca un rol crucial în această extindere a rețelei de interacțiune genetică. Un număr substanțial al acestor molecule a apărut în momentul achiziției lamelocitelor, ele sunt exprimate la momentul potrivit în răspunsul imun și prezintă semnături puternice de selecție pozitivă (Wertheim et al., 2005; Salazar-Jaramillo et al., 2014).
De asemenea, în cadrul unei singure specii, D. melanogaster, răspunsurile imune prezintă o mare variație genetică. Populațiile de câmp colectate din întreaga Europă prezintă diferențe substanțiale în ceea ce privește capacitatea de a încapsula cu succes ouăle de parazitoizi (Kraaijeveld și van Alphen, 1995; Kraaijeveld și Godfray, 1999; Gerritsma et al., 2013). Aparent, costurile și beneficiile unei apărări imunitare puternice diferă din punct de vedere geografic, ceea ce duce la modularea și diferențierea rețelelor genetice co-adaptate. Acest lucru s-a reflectat, de asemenea, în răspunsul hemocitar după atacul parazitului. Liniile de câmp au variat considerabil în ceea ce privește numărul absolut și relativ al diferitelor hemocite pe care le-au produs ca răspuns la atacul parazitoidelor, chiar și în rândul liniilor care au avut un mare succes la încapsulare (Gerritsma et al., 2013). Acest lucru subliniază din nou faptul că fondul genetic al unei populații și presiunile de selecție locale combinate conduc la răspunsuri evolutive alternative. Compararea genomurilor indivizilor rezistenți și susceptibili din mai multe populații poate dezvălui variația adaptativă în arhitectura genetică a acestei trăsături.
Pentru a cartografia modificările din genom în timpul evoluției rezistenței crescute, am efectuat o evoluție experimentală pentru rezistența la parazitoizi. În laborator, am expus o populație mare de animale de rasă la parazitoizi. Doar larvele care au reușit să supraviețuiască atacului parazitoidelor au fost lăsate să contribuie la generația următoare. Cu această abordare, am crescut nivelul de rezistență de la 20 la ∼50% din larvele care au supraviețuit atacului parazitoidelor după numai cinci generații de selecție. Atunci când am măsurat modificările în expresia genelor în populațiile selectate, în comparație cu expresia genelor din liniile de control, chiar înainte de atacul parazitoidelor, am descoperit câteva sute de gene care erau ușor reglementate diferențiat (Wertheim et al., 2011). Modificările au implicat mai ales gene care nu au fost exprimate diferențiat în timpul răspunsului imunitar, ceea ce indică faptul că modificările evolutive nu au pre-activat răspunsul imunitar în anticiparea atacului parazitoidelor, ci a modulat calea de dezvoltare canonică, ceea ce a condus (de asemenea) la o creștere a capacităților sale defensive. Am repetat acest experiment, iar apoi am secvențiat genomurile populațiilor selectate și de control. În genomurile liniilor care au evoluat cu o rezistență sporită, am găsit semnături de selecție pe mai multe regiuni îngust definite ale genomului (Jalvingh et al., 2014). Unele dintre aceste regiuni s-au suprapus, de asemenea, cu regiunile care au prezentat modificări ale expresiei după selecția pentru rezistență sporită (Wertheim et al., 2011; Jalvingh et al., 2014). Astfel, o măturare selectivă rapidă și puternică asupra unei trăsături complexe precum imunitatea poate afecta în continuare regiuni genomice multiple, dar foarte localizate.
Provocări viitoare
Cum vom împăca schimbările evolutive pe termen lung, cum ar fi achiziția de noi gene în rețelele de interacțiune genetică, și schimbările evolutive pe termen scurt, cum ar fi variantele de secvență care pot fi măturate într-o populație? Cheia în acest sens este (i) reconstruirea rețelei de interacțiune genetică care stă la baza trăsăturilor complexe și (ii) caracterizarea rolului variației genetice în cadrul acestor rețele. Rețelele genetice se pot extinde cu noi gene prin, de exemplu, duplicări, pot deveni interconectate cu alte rețele sau module, iar mici variații de secvență pot modula activitatea și topologia rețelelor. Dacă putem descompune rețelele genetice și evalua rolul variantelor genetice într-un context de rețea, acest lucru ne va permite, în cele din urmă, să determinăm modul în care variația genetică se traduce în variație fenotipică. Acest lucru ne va îmbunătăți, de asemenea, înțelegerea bazei moleculare a bolilor umane complexe și a evoluției imunității înnăscute și dobândite (Cooper și Alder, 2006; Manolio et al., 2009; Star et al., 2011; Mackay și Moore, 2014; Sironi et al., 2015).
Abordările biologice ale sistemelor vor fi de neprețuit pentru a desluși rețelele complexe de interacțiune genetică. Acolo, se dezvoltă modele matematice pentru a descrie mecanismele moleculare care stau la baza unei trăsături și pentru a prezice dinamica grupurilor de componente ale rețelei care interacționează. Modelele se bazează pe date de genetică moleculară și genomică. În prezent, biologia sistemelor se aplică în principal la trăsături specifice la organismele unicelulare, iar aceasta este considerată limita a ceea ce se poate realiza (Papp et al., 2011). Cu toate acestea, este probabil ca acest model să nu fie reprezentativ pentru evoluția în cazul organismelor multicelulare cu reproducere sexuată. În loc să așteptăm ca modelele simplificate să reflecte cu exactitate sub-rețelele mici, trebuie să dezvoltăm și să ne perfecționăm metodele pentru a utiliza și cuantifica proprietățile emergente din marea cantitate de date genomice. Putem deduce rețelele de interacțiune genetică din matrici de corelație sau de coexpresie a proteinelor-proteine sau a transcriptelor (Shannon et al., 2003; Langfelder și Horvath, 2008) și le putem îmbina cu analize ale variantelor naturale (Nuzhdin et al., 2012). Ar trebui să dezvoltăm în continuare aceste metode și abordări alternative pentru a exploata pe deplin măsurătorile noastre asupra datelor genomice și pentru a converti aceste măsurători cantitative în analize de rețea. Deși datele genomice în sine nu vor oferi un răspuns complet la ceea ce determină capacitatea de adaptare a vieții, ele ne permit să cuantificăm și să observăm ceea ce se întâmplă la nivel molecular în timpul evoluției. Atunci când combinăm și integrăm acest lucru cu eterogenitatea mediului ca motor al adaptărilor, am putea fi capabili să dezvăluim mecanismele moleculare complexe de adaptare și evoluție.
Declarație privind conflictul de interese
Autorul declară că cercetarea a fost efectuată în absența oricăror relații comerciale sau financiare care ar putea fi interpretate ca un potențial conflict de interese.
Recunoștințe
Le mulțumesc lui Kirsten Jalvingh, Laura Salazar-Jaramillo și Sylvia Gerritsma pentru discuțiile valoroase. BW a beneficiat de finanțare din partea Organizației Olandeze pentru Cercetare Științifică (NWO; grant 864.08.008).
Aggarwal, K., și Silverman, N. (2008). Reglarea pozitivă și negativă a răspunsului imunitar al Drosophilei. BMB Rep. 41, 267-277. doi: 10.5483/BMBRep.2008.41.4.267
PubMed Abstract | CrossRef Full Text | Google Scholar
Ayroles, J. F., Carbone, M. A., Stone, E. A., Jordan, K. W., Lyman, R. F., Magwire, M. M., et al. (2009). Genetica sistemelor de trăsături complexe la Drosophila melanogaster. Nat. Genet. 41, 299-307. doi: 10.1038/ng.332
PubMed Abstract |Ref Full Text | Google Scholar
Brennan, C. A., și Anderson, K. V. (2004). Drosophila: genetica recunoașterii și răspunsului imunitar înnăscut. Annu. Rev. Immunol. 22, 457-483. doi: 10.1146/annurev.immunol.22.012703.104626
PubMed Abstract | CrossRef Full Text | Google Scholar
Buchmann, K. (2014). Evoluția imunității înnăscute: indicii de la nevertebrate, prin intermediul peștilor, la mamifere. Front. Immunol. 5:459. doi: 10.3389/fimmu.2014.00459
PubMed Abstract | Text integral | Google Scholar
Carpenter, S., Ricci, E. P., Mercier, B. C., Moore, M. J., și Fitzgerald, K. A. (2014). Reglarea post-transcripțională a expresiei genice în imunitatea înnăscută. Nat. Rev. Immunol. 14, 361-376. doi: 10.1038/nri3682
PubMed Abstract | Textul integral | Google Scholar
Cooper, M. D., și Alder, M. N. (2006). Evoluția sistemelor imunitare adaptative. Cell 124, 815-822. doi: 10.1016/j.cell.2006.02.001
PubMed Abstract | Text integral | Google Scholar
Dobzhansky, T. (1964). Biologie, moleculară și organismică. Am. Zool. 4, 443-452.
PubMed Abstract | Google Scholar
Drosophila 12 Genomes Consortium, Clark, A. G., Eisen, M. B., Smith, D. R., Bergman, C. M., Oliver, B., et al. (2007). Evoluția genelor și a genomurilor în filogenia Drosophila. Nature 450, 203-218. doi: 10.1038/nature06341
PubMed Abstract | Text integral | Google Scholar
Edwards, A. C., Ayroles, J. F., Stone, E. A., Carbone, M. A., Lyman, R. F., și Mackay, T. F. C. (2009). O rețea transcripțională asociată cu variația naturală în comportamentul agresiv al Drosophilei. Genome Biol. 10, R76. doi: 10.1186/gb-2009-10-7-r76
PubMed Abstract | Text integral | Google Scholar
Eslin, P., și Doury, G. (2006). Musca Drosophila subobscura: un caz natural de deficiență a imunității înnăscute. Dev. Comp. Immunol. 30, 977-983. doi: 10.1016/j.dci.2006.02.007
PubMed Abstract | Full CrossRef Text | Google Scholar
Evans, C. J., Hartenstein, V., și Banerjee, U. (2003). Mai gros decât sângele: mecanisme conservate în Drosophila și hematopoieza vertebratelor. Dev. Cell 5, 673-690. doi: 10.1016/S1534-5807(03)00335-6
PubMed Abstract | CrossRef Full Text | Google Scholar
Fay, J. C., McCullough, H. L., Sniegowski, P. D., și Eisen, M. B. (2004). Variația genetică a populației în expresia genelor este asociată cu variația fenotipică la Saccharomyces cerevisiae. Genome Biol. 5, R26. doi: 10.1186/gb-2004-5-4-4-r26
PubMed Abstract | CrossRef Full Text | Google Scholar
Fleury, F., Gibert, P., Ris, N., și Allemand, R. (2009). Ecology and life history evolution of frugivorous Drosophila parasitoids. Adv. Parasitol. 70, 3-44. doi: 10.1016/S0065-308X(09)70001-6
PubMed Abstract | Full CrossRef Text | Google Scholar
Gasch, A. P., Spellman, P. T., Kao, C. M., Carmel-Harel, O., Eisen, M. B., Storz, G., et al. (2000). Programe de expresie genomică în răspunsul celulelor de drojdie la schimbările de mediu. Mol. Biol. Cell 11, 4241-4257. doi: 10.1091/mbc.11.12.4241
PubMed Abstract | CrossRef Full Text | Google Scholar
Gerritsma, S., de Haan, A., van de Zande, L., și Wertheim, B. (2013). Variația naturală a hemocitelor diferențiate este legată de rezistența la parazitoizi la Drosophila melanogaster. J. Insect Physiol. 59, 148-158. doi: 10.1016/j.jinsphys.2012.09.017
PubMed Abstract | CrossRef Full Text | Google Scholar
Havard, S., Doury, G., Ravallec, M., Brehélin, M., Prevost, G., și Eslin, P. (2012). Caracterizarea structurală și funcțională a pseudopodocitelor, o celulă imunitară zbârcită produsă de două specii de Drosophila din grupul obscura. Dev. Comp. Immunol. 36, 323-331. doi: 10.1016/j.dci.2011.05.009
PubMed Abstract | Text integral | Google Scholar
Hunt, P., Martinelli, A., Modrzynska, K., Borges, S., Creasey, A., Rodrigues, L., et al. (2010). Evoluția experimentală, analiza genetică și resecvențierea genomului dezvăluie mutația care conferă rezistență la artemisinină într-o linie izogenă de paraziți de malarie. BMC Genomics 11:499. doi: 10.1186/1471-2164-11-499
PubMed Abstract | Full CrossRef Text | Google Scholar
Ivanov, P., și Anderson, P. (2013). Rețele de reglementare post-transcripțională în imunitate. Immunol. Rev. 253, 253-272. doi: 10.1111/imr.12051
PubMed Abstract | Full Text | Google Scholar
Jalvingh, K. M., Chang, P. L., Nuzhdin, S. V., și Wertheim, B. (2014). Modificări genomice în condiții de evoluție rapidă: selecția pentru rezistența la parazitoizi. Proc. Biol. Sci. 281, 20132303. doi: 10.1098/rspbb.2013.2303
PubMed Abstract | CrossRef Full Text | Google Scholar
Jones, F. C., Grabherr, M. G., Chan, Y. F. F., Russell, P., Mauceli, E., Johnson, J., et al. (2012). Baza genomică a evoluției adaptative în cazul stickleback-urilor threespine. Nature 484, 55-61. doi: 10.1038/nature10944
PubMed Abstract | Textul integral | Google Scholar
Knight, C. G., Zitzmann, N., Prabhakar, S., Antrobus, R., Dwek, R., Hebestreit, H., et al. (2006). Unraveling adaptive evolution: how a single point mutation affects the protein coregulation network. Nat. Genet. 38, 1015. doi: 10.1038/ng1867
PubMed Abstract |Ref Full Text | Google Scholar
Kraaijeveld, A. R., și Godfray, H. C. J. (1999). Modele geografice în evoluția rezistenței și virulenței la Drosophila și la parazitoizii săi. Am. Nat. 153, S61-S74. doi: 10.1086/303212
CrossRef Full Text | Google Scholar
Kraaijeveld, A. R., Ferrari, J., și Godfray, H. C. J. (2002). Costurile rezistenței în interacțiunile insecte-paraziți și insecte-parazitoizi. Parasitology 125(Suppl.), S71-S82. doi: 10.1017/S003118/S0031182002001750
PubMed Abstract | CrossRef Full Text | Google Scholar
Kraaijeveld, A. R., and van Alphen, J. J. M. (1995). Variația geografică a capacității de încapsulare a larvelor de Drosophila melanogaster și dovezi pentru componentele specifice parazitoidelor. Evol. ecol. 9, 10-17. doi: 10.1007/BF01237692
CrossRef Full Text | Google Scholar
Langfelder, P., și Horvath, S. (2008). WGCNA: un pachet R pentru analiza rețelelor de corelație ponderată. BMC Bioinformatics 9:559. doi: 10.1186/1471-2105-9-559
PubMed Abstract | Ref. Full Text | Google Scholar
Lavine, M. D., și Strand, M. R. (2002). Hemocitele insectelor și rolul lor în imunitate. Insect Biochem. Mol. Biol. 32, 1295-1309. doi: 10.1016/S0965-1748(02)00092-9
PubMed Abstract | CrossRef Full Text | Google Scholar
Lazzaro, B. P., and Little, T. J. (2009). Imunitatea într-o lume variabilă. Philos. Trans. R. Soc. Lond. B Biol. Sci. 364, 15-26. doi: 10.1098/rstb.2008.0141
PubMed Abstract | Refef Full Text | Google Scholar
Lazzaro, B. P., și Schneider, D. S. (2014). Genetica imunității. G3 (Bethesda) 4, 943-945. doi: 10.1534/g3.114.011684
PubMed Abstract | CrossRef Full Text | Google Scholar
Lehner, B. (2013). De la genotip la fenotip: lecții de la organismele model pentru genetica umană. Nat. Rev. Genet. 14, 168-178. doi: 10.1038/nrg3404
PubMed Abstract |Ref Full Text | Google Scholar
Lemaitre, B., și Hoffmann, J. (2007). Apărarea gazdă a Drosophila melanogaster. Annu. Rev. Immunol. 25, 697-743. doi: 10.1146/annurev.immunol.25.022106.141615
PubMed Abstract | CrossRef Full Text | Google Scholar
Liew, F. Y., Xu, D., Brint, E. K., și O’Neill, L. A. J. (2005). Reglarea negativă a răspunsurilor imune mediate de receptorii Toll-like. Nat. Rev. Immunol. 5, 446-458. doi: 10.1038/nri1630
PubMed Abstract | Ref. Full Text | Google Scholar
Linnen, C. R., Poh, Y.-P., Peterson, B. K., Barrett, R. D. H., Larson, J. G., Jensen, J. D., et al. (2013). Evoluția adaptivă a mai multor trăsături prin mutații multiple la o singură genă. Science 339, 1312-1316. doi: 10.1126/science.1233213
PubMed Abstract | Text integral | Google Scholar
Mackay, T. F., și Moore, J. H. (2014). De ce epistaza este importantă pentru abordarea geneticii complexe a bolilor umane complexe. Genome Med. 6, 42. doi: 10.1186/gm561
PubMed Abstract | Full CrossRef Full Text | Google Scholar
Manolio, T. A., Collins, F. S., Cox, N. J., Goldstein, D. B., Hindorff, L. A., Hunter, D. J., et al. (2009). Găsirea heritabilității lipsă a bolilor complexe. Nature 461, 747-753. doi: 10.1038/nature08494
PubMed Abstract | Full Text | Google Scholar
Márkus, R., Lerner, Z., Honti, V., Csordás, G., Zsámboki, J., Cinege, G., et al. (2015). Hemocitele gigantice multinucleate sunt celule efectoare în răspunsurile imune mediate de celule ale Drosophilei. J. Innate Immun. doi: 10.1159/000369618 .
PubMed Abstract | Full CrossRef Text | Google Scholar
Moore, J. H., și Williams, S. M. (eds). (2015). Epistasis. New York, NY: Springer. doi: 10.1007/978-1-4939-2155-3
CrossRef Full Text | Google Scholar
Nagy, A., Perrimon, N., Sandmeyer, S., și Plasterk, R. (2003). Adaptarea genomului: puterea abordărilor genetice. Nat. Genet. 33(Suppl.), 276-284. doi: 10.1038/ng1115
PubMed Abstract | Text integral | Google Scholar
Nowick, K., Gernat, T., Almaas, E., și Stubbs, L. (2009). Diferențele dintre modelele de expresie a genelor umane și cele ale cimpanzeilor definesc o rețea evolutivă de factori de transcripție în creier. Proc. Natl. acad. Sci. U.S.A. 106, 22358-22363. doi: 10.1073/pnas.0911376106
PubMed Abstract | CrossRef Full Text | Google Scholar
Nuzhdin, S. V., Friesen, M. L., și McIntyre, L. M. (2012). Cartografierea genotipurilor-fenotipurilor într-o lume post-GWAS. Trends Genet. 28, 421-426. doi: 10.1016/j.tig.2012.06.003
PubMed Abstract | CrossRef Full Text | Google Scholar
Obbard, D. J., Welch, J. J., J. J., Kim, K.-W., și Jiggins, F. M. (2009). Cuantificarea evoluției adaptative în sistemul imunitar al Drosophilei. PLoS Genet. 5:e1000698. doi: 10.1371/journal.pgen.1000698
PubMed Abstract | Text integral | Google Scholar
Papp, B., Notebaart, R. A., și Pál, C. (2011). Systems-biology approaches for predicting genomic evolution (Abordări de biologie sistemică pentru predicția evoluției genomice). Nat. Rev. Genet. 12, 591-602. doi: 10.1038/nrg3033
PubMed Abstract |Ref Full Text | Google Scholar
Pedra, J. H. F., McIntyre, L. M., Scharf, M. E., și Pittendrigh, B. R. (2004). Genome-wide transcription profile of field- and laboratory-selected dichlorodiphenyltrichloroethane (DDT)-resistant Drosophila. Proc. Natl. acad. Sci. U.S.A. 101, 7034-7039. doi: 10.1073/pnas.040058010101
PubMed Abstract | CrossRef Full Text | Google Scholar
Phillips, P. C. (2008). Epistasis-rolul esențial al interacțiunilor genice în structura și evoluția sistemelor genetice. Nat. Rev. Genet. 9, 855-867. doi: 10.1038/nrg2452
PubMed Abstract |Ref Full Text | Google Scholar
Rolff, J., și Siva-Jothy, M. T. (2003). Imunologia ecologică a nevertebratelor. Science 301, 472-475. doi: 10.1126/science.1080623
PubMed Abstract | Text integral | Google Scholar
Sackton, T. B., Lazzaro, B. P., Schlenke, T. A., Evans, J. D., Hultmark, D., și Clark, A. G. (2007). Evoluția dinamică a sistemului imunitar înnăscut la Drosophila. Nat. Genet. 39, 1461-1468. doi: 10.1038/ng.2007.60
PubMed Abstract | Full Text | Google Scholar
Salazar-Jaramillo, L., Paspati, A., van de Zande, L., Vermeulen, C. J., Schwander, T., și Wertheim, B. (2014). Evoluția unui răspuns imun celular la Drosophila: o analiză comparativă fenotipică și genomică. Genome Biol. Evol. 6, 273-289. doi: 10.1093/gbe/evu012
PubMed Abstract | Text integral | Google Scholar
Schadt, E. E. (2009). Rețelele moleculare ca senzori și motoare ale bolilor umane comune. Nature 461, 218-223. doi: 10.1038/nature08454
PubMed Abstract | Text integral | Google Scholar
Schlenke, T. A., Morales, J., Govind, S., și Clark, A. G. (2007). Strategii de infecție contrastante la viespile parazitoide generaliste și specializate de Drosophila melanogaster. PLoS Pathog. 3:e158. doi: 10.1371/journal.ppat.0030158
PubMed Abstract | Full Text | Google Scholar
Schmid-Hempel, P. (2003). Variația în apărarea imunitară ca o chestiune de ecologie evolutivă. Proc. R. Soc. B Biol. Sci. 270, 357-366. doi: 10.1098/rspbb.2002.2265
PubMed Abstract | Text integral | Google Scholar
Schmid-Hempel, P. (2005). Ecologia evolutivă a apărării imunitare a insectelor. Annu. Rev. Entomol. 50, 529-551. doi: 10.1146/annurev.ento.50.071803.130420
PubMed Abstract | CrossRef Full Text | Google Scholar
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: un mediu software pentru modele integrate de rețele de interacțiune biomoleculară. Genome Res. 13, 2498-2504. doi: 10.1101/gr.1239303
PubMed Abstract | Full CrossRef Text | Google Scholar
Silverman, N., și Maniatis, T. (2001). Căile de semnalizare NF-κB în imunitatea înnăscută a mamiferelor și insectelor. Genes Dev. 15, 2321-2342. doi: 10.1101/gad.909001
PubMed Abstract | Refef Full Text | Google Scholar
Sironi, M., Cagliani, R., Forni, D., și Clerici, M. (2015). Perspective evolutive în interacțiunile gazdă-patogen din datele de secvență ale mamiferelor. Nat. Rev. Genet. 16, 224-236. doi: 10.1038/nrg3905
PubMed Abstract |Ref Full Text | Google Scholar
Stapley, J., Reger, J., Feulner, P. G. D., Smadja, C., Galindo, J., Ekblom, R., et al. (2010). Genomica adaptării: următoarea generație. Trends Ecol. Evol. 25, 705-712. doi: 10.1016/j.tree.2010.09.002
PubMed Abstract | CrossRef Full Text | Google Scholar
Star, B., Nederbragt, A. J., Jentoft, S., Grimholt, U., Malmstrøm, M., Gregers, T. F., et al. (2011). Secvența genomului codului de Atlantic dezvăluie un sistem imunitar unic. Nature 477, 207-210. doi: 10.1038/nature10342
PubMed Abstract | Full Text | Google Scholar
Stearns, F. W. (2010). O sută de ani de pleiotropie: o retrospectivă. Genetica 186, 767-773. doi: 10.1534/genetics.110.122549
PubMed Abstract | Text integral | Google Scholar
Storz, J. F., și Wheat, C. W. (2010). Integrarea abordărilor evolutive și funcționale pentru a deduce adaptarea la loci specifici. Evolution 64, 2489-2509. doi: 10.1111/j.1558-5646.2010.01044.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Tenaillon, O., Rodríguez-Verdugo, A., Gaut, R. L., McDonald, P., Bennett, A. F., Long, A. D., et al. (2012). Diversitatea moleculară a convergenței adaptative. Science 335, 457-461. doi: 10.1126/science.1212986
PubMed Abstract | Full Text | Google Scholar
Turner, T. L., Stewart, A. D., Fields, A. T., Rice, W. R., și Tarone, A. M. (2011). Resecvențierea pe bază de populație a populațiilor evoluate experimental dezvăluie baza genetică a variației dimensiunii corporale la Drosophila melanogaster. PLoS Genet. 7:e1001336. doi: 10.1371/journal.pgen.1001336
PubMed Abstract | Full CrossRef Text | Google Scholar
Wagner, G. P., și Altenberg, L. (1996). Adaptări complexe și evoluția evoluției evolutivității. Evolution 50, 967-976. doi: 10.2307/2410639
CrossRef Full Text | Google Scholar
Wertheim, B., Kraaijeveld, A. R., Hopkins, M. G., Walther Boer, M., și Godfray, H. C. (2011). Genomica funcțională a evoluției rezistenței crescute la parazitism la Drosophila. Mol. Ecol. 20, 932-949. doi: 10.1111/j.1365-294X.2010.04911.x
PubMed Abstract | CrossRef Full Text | Google Scholar
Wertheim, B., Kraaijeveld, A. R., Schuster, E., Blanc, E., Hopkins, M., Pletcher, S. D., et al. (2005). Expresia genelor la nivelul genomului ca răspuns la atacul parazitului la Drosophila. Genome Biol. 11, R94. doi: 10.1186/gb-2005-6-11-r94
PubMed Abstract | Full CrossRef Text | Google Scholar
Zak, D. E., Tam, V. C. și Aderem, A. (2014). Analiza la nivel de sistem a imunității înnăscute. Annu. Rev. Immunol. 32, 547-577. doi: 10.1146/annurev-immunol-032713-120254
PubMed Abstract | CrossRef Full Text | Google Scholar
.