DNAポリメラーゼはどのように突然変異を防ぐのか
突然変異は、特定の生物の塩基配列が永久に変化することです。 DNA複製の誤りや外部からの変異原によって生じることがあります。 突然変異の影響は、細胞にとって有益なものと有害なもののどちらにもなり得る。 しかし、細胞は突然変異を防ぐために様々な種類のメカニズムを持っています。 DNAの複製に関与する酵素であるDNAポリメラーゼは、DNA複製の際のエラーを防ぐためのいくつかの機構を備えている。 DNA複製の際には、ミスペアとなった塩基はプルーフリーディングによって置き換えられる。 DNA複製の直後には、残ったミスペア塩基が鎖指向性ミスマッチ修復により置換される。 さらに、外的要因によって生じた変異は、切除修復、化学的逆転、二本鎖切断修復などいくつかのメカニズムによって修復される。 もし、損傷が可逆的であれば、細胞はアポトーシスを起こし、子孫に欠陥のあるDNAを受け継がないようにする。
Key Areas Covered
1. 突然変異とは何か
– 定義、種類、原因
2.DNAポリメラーゼはどのようにして突然変異を防いでいるのか
– 校正、鎖指向性ミスマッチ修復
主要用語。 DNAポリメラーゼ、鎖状指向性ミスマッチ修復、Mutタンパク質、突然変異、校正

What is a Mutation
突然変異とは、ゲノム上の塩基配列に永久的かつ遺伝性の変化が起こることで、この変化は遺伝的な変異を意味します。 突然変異は、DNA複製のエラーや突然変異原として知られる外的要因によって生じることがある。
点突然変異
点突然変異は、一塩基の置換で、その変異の形態は、点突然変異、フレームシフト突然変異、染色体突然変異の3つがある。 点変異にはミスセンス変異、ナンセンス変異、サイレント変異の3種類がある。 ミスセンス変異は遺伝子の単一コドンを変化させ、ポリペプチド鎖のアミノ酸を変化させる。 ナンセンス変異はコドンの配列が変わるものの、アミノ酸の配列は変化しない。 サイレント変異は、単一のコドンを同じアミノ酸を表す別のコドンに変更する。 点突然変異は、DNA複製のエラーや突然変異誘発物質によって引き起こされる。 図1
図1:点突然変異
フレームシフト突然変異
フレームシフト突然変異は、ゲノムに単一または複数のヌクレオチドが挿入または欠落する突然変異である。 挿入、欠失、重複の3つがフレームシフト変異の種類である。 挿入は配列に1つまたは数個のヌクレオチドが追加されることであり、欠失は配列から数個のヌクレオチドが削除されることである。 重複は、数個のヌクレオチドが繰り返されることである。
染色体突然変異
染色体突然変異は、染色体の一部が変化することです。 染色体突然変異には、転座、遺伝子重複、染色体内欠失、逆位、ヘテロ接合性喪失があります。 転座は、非相同染色体間で染色体の一部が交換されることです。 遺伝子重複では、特定の対立遺伝子のコピーが複数出現し、遺伝子量が増加することがある。 染色体の一部が欠けることを染色体内欠失といいます。 逆位は、染色体の向きを変える。 遺伝子のヘテロ接合性は、欠失や遺伝子組換えにより、一方の染色体の対立遺伝子が失われることにより、失われることがある。 染色体の突然変異は、主に外部からの変異原やDNAの機械的損傷によって起こります。
DNAポリメラーゼはどのようにして突然変異を防ぐのか
DNAポリメラーゼは、DNA複製の際に成長する鎖に塩基を付加する役割を果たす酵素で、この酵素の働きによって突然変異が起こります。 ゲノムの塩基配列は、特定の生物の発生と機能を決定するため、DNA複製の際に、既存のゲノムの正確な複製を合成することが不可欠です。 一般に、DNAポリメラーゼはDNA複製の際に高い忠実度を保ち、109個のヌクレオチドを付加する際に1個のミスマッチを取り込むだけである。 したがって、標準的な相補的塩基対に加えて窒素塩基の間にも不一致が生じると、DNAポリメラーゼはそのヌクレオチドを成長する鎖に付加し、頻繁に突然変異を生じさせることになる。
プルーフリーディング
プルーフリーディングとは、成長するDNA鎖から誤った塩基対を修正する最初のメカニズムを指し、DNAポリメラーゼによって実行される。 DNAポリメラーゼは2つのステップでプルーフリーディングを行う。 最初のプルーフリーディングは、成長する鎖に新しいヌクレオチドが付加される直前に行われる。 DNAポリメラーゼに対する正しいヌクレオチドの親和力は、正しくないヌクレオチドの親和力の何倍も高いのです。 しかし、DNAポリメラーゼの作用でヌクレオチドが成長中の鎖に結合する前に、酵素は水素結合によって鋳型に結合した直後に立体構造を変化させるはずである。 塩基対が正しくないヌクレオチドは、DNAポリメラーゼの構造変化の際に鋳型から解離しやすくなっている。 そこで、このステップでは、DNAポリメラーゼがヌクレオチドを成長中の鎖に永久に付加する前に「ダブルチェック」することができる。 DNAポリメラーゼの校正機構を図2に示す。
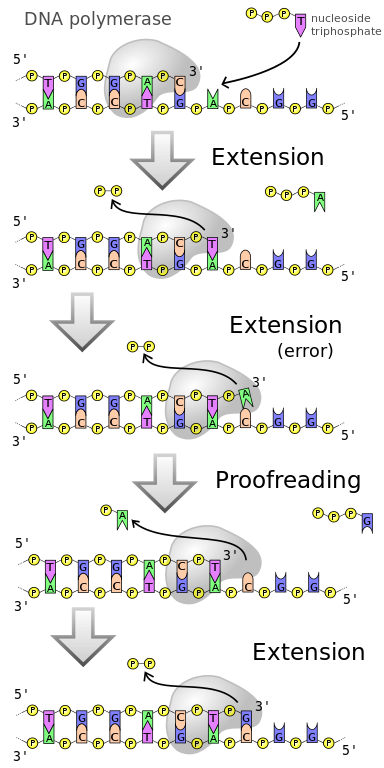
Figure 2: Proofreading
2番目の校正ステップはexonucleolytic proofreadingと呼ばれ、核酸分解による校正を行う。 これは、まれに、成長中の鎖にミスマッチのヌクレオチドが組み込まれた直後に起こる。 DNAポリメラーゼは、ミスマッチのあるヌクレオチドの隣に2番目のヌクレオチドを付加することができない。
Strand-Directed Mismatch Repair
校正メカニズムにもかかわらず、DNAポリメラーゼはDNA複製の際に成長する鎖に誤ったヌクレオチドを組み込むことがある。 プルーフリーディングを逃れた複製エラーは、鎖指向性ミスマッチ修復によって除去される。 このシステムは、ミスマッチした塩基対に起因するDNAらせんの歪みポテンシャルを検出する。 しかし、修復システムは、ミスマッチを置き換える前に、既存の塩基から誤った塩基を特定する必要がある。 一般に、大腸菌は、新しく合成されたDNA鎖がすぐにDNAメチル化を受けない可能性があるため、DNAメチル化システムに依存して、二重らせん内の古いDNA鎖を認識することができる。 大腸菌では、GATCのA残基がメチル化されている。 大腸菌では、GATCのA残基がメチル化され、鎖指向性ミスマッチ修復システムの働きにより、DNA複製の忠実度はさらに102倍向上する。 真核生物、細菌、大腸菌におけるDNAミスマッチ修復経路を図3に示す。
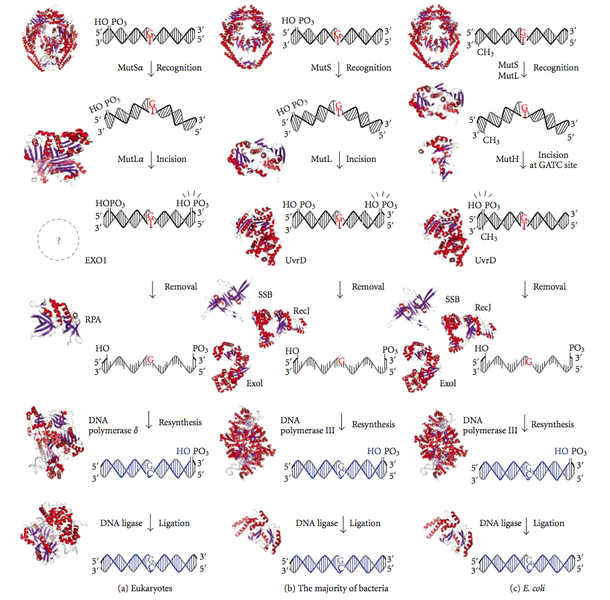
図3:真核生物、細菌、大腸菌におけるDNAミスマッチ修復
鎖指向ミスマッチ修復では3つの複合タンパク質が新しく合成されたDNA鎖を移動しながら修復している。 MutSとして知られる最初のタンパク質は、DNA二重らせんのゆがみを検知して結合する。 MutLと呼ばれる2番目のタンパク質はMutSを検出して結合し、MutHと呼ばれる3番目のタンパク質はメチル化されていない鎖と新しく合成された鎖を区別して引きつける。 MutHは、メチル化されていないDNA鎖を、GATC配列のG残基のすぐ上流で切断し、メチル化されたDNA鎖を合成する。 ミスマッチの下流側の鎖は、エキソヌクレアーゼが分解を担当する。 しかし、このシステムでは、DNAポリメラーゼ1によって容易に再合成される10塩基以下の領域は分解されない。 真核生物のMutタンパク質は大腸菌のものと相同性がある。
結論
突然変異とは、DNA複製の誤りや外部の変異原の影響により、ゲノムのヌクレオチド配列が永久的に変化することである。 DNA複製のエラーは、プルーフリーディングと鎖指向性ミスマッチ修復という2つのメカニズムによって修正することができる。 プルーフリーディングは、DNA合成の際にDNAポリメラーゼ自身が行うものである。 鎖指向性ミスマッチ修復は、DNA複製直後にMutタンパク質によって行われる。 しかし、これらの修復機構はゲノムの完全性の維持に関与している
参考文献:
1. Alberts, Bruce. “DNA複製機構”. 細胞の分子生物学. 4th edition., U.S. National Library of Medicine, 1 Jan. 1970, Available here.
2. Brown, Terence A. “Mutation, Repair and Recombination”(突然変異、修復、組み換え). ゲノム. 第2版, U.S. National Library of Medicine, 1 Jan. 1970, Available here.
Image Courtesy:
1. “Different Types of Mutations” By Jonsta247 – This file was derived from:Point mutations-en.png (GFDL) via Commons Wikimedia
2. “DNA polymerase” By I, Madprime (CC BY-SA 3.0) via Commons Wikimedia
3. “DNA mismatch repair” By Kenji Fuki – (CC BY 3.0) via Commons Wikimedia
![]()