Comment l’ADN polymérase prévient-il les mutations
Les mutations sont des modifications permanentes de la séquence nucléotidique d’un organisme particulier. Elles peuvent survenir en raison des erreurs de réplication de l’ADN ou de mutagènes externes. L’effet d’une mutation peut être bénéfique ou délétère pour la cellule. Toutefois, les cellules mettent en œuvre divers types de mécanismes pour prévenir les mutations. L’ADN polymérase, qui est l’enzyme impliquée dans la réplication de l’ADN, est dotée de plusieurs mécanismes pour prévenir les erreurs lors de la réplication de l’ADN. Pendant la réplication de l’ADN, les bases mal appariées sont remplacées par la relecture. Immédiatement après la réplication de l’ADN, les bases altérées restantes sont remplacées par la réparation des mésappariements dirigée par le brin. En outre, les mutations causées par des facteurs externes sont réparées par plusieurs mécanismes tels que la réparation par excision, l’inversion chimique et la réparation des cassures double-brin. Si le dommage est réversible, la cellule est soumise à l’apoptose afin d’éviter de transmettre l’ADN défectueux à la descendance.
Domaines clés couverts
1. Qu’est-ce qu’une mutation
– Définition, types, causes
2. Comment l’ADN polymérase prévient-elle les mutations
– Relecture, réparation des mésappariements dirigée par le brin
Termes clés : ADN polymérase, réparation des mésappariements dirigée par brin, protéines mut, mutation, relecture

Qu’est-ce qu’une mutation
Une mutation désigne un changement permanent et héréditaire de la séquence nucléotidique du génome. Les mutations peuvent survenir en raison des erreurs de réplication de l’ADN ou de facteurs externes appelés mutagènes. Les trois formes de mutations sont les mutations ponctuelles, les mutations par décalage de cadre et les mutations chromosomiques.
Mutations ponctuelles
Les mutations ponctuelles sont des substitutions d’un seul nucléotide. Les trois types de mutations ponctuelles sont les mutations faux-sens, non-sens et silencieuses. La mutation faux-sens modifie un seul codon du gène, altérant l’acide aminé de la chaîne polypeptidique. Les mutations non-sens modifient la séquence des codons, mais pas celle des acides aminés. Les mutations silencieuses modifient un codon unique en un autre codon qui représente le même acide aminé. Les mutations ponctuelles sont causées par des erreurs de réplication de l’ADN et par des agents mutagènes. Différents types de mutations ponctuelles sont présentés dans la figure 1.
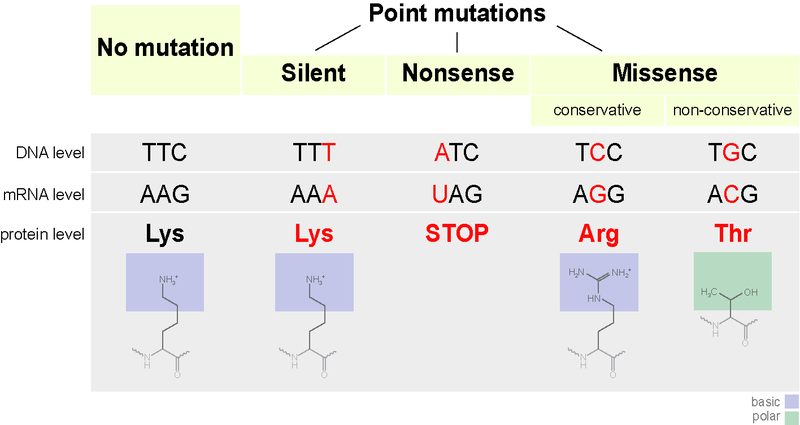
Figure 1 : Mutations ponctuelles
Mutations par décalage de cadre
Les mutations par décalage de cadre sont des insertions ou des délétions d’un ou plusieurs nucléotides du génome. Les insertions, les délétions et les duplications sont les trois types de mutations de décalage de cadre. Les insertions sont l’ajout d’un ou plusieurs nucléotides à la séquence, tandis que les délétions sont la suppression de plusieurs nucléotides de la séquence. Les duplications sont la répétition de plusieurs nucléotides. Les mutations de type Frameshift sont également causées par des erreurs de réplication de l’ADN et par des agents mutagènes.
Mutations chromosomiques
Les mutations chromosomiques sont des altérations de segments de chromosomes. Les types de mutations chromosomiques sont les translocations, les duplications de gènes, les délétions intra-chromosomiques, les inversions et la perte d’hétérozygotie. Les translocations sont les échanges de parties de chromosomes entre des chromosomes non homologues. Dans le cas de la duplication de gènes, de multiples copies d’un allèle particulier peuvent apparaître, augmentant ainsi le dosage du gène. Les suppressions de segments de chromosomes sont appelées délétions intra-chromosomiques. Les inversions modifient l’orientation d’un segment de chromosome. L’hétérozygotie d’un gène peut être perdue en raison de la perte d’un allèle dans un chromosome par délétion ou recombinaison génétique. Les mutations chromosomiques sont principalement causées par des mutagènes externes et en raison de dommages mécaniques à l’ADN.
Comment l’ADN polymérase prévient-elle les mutations
L’ADN polymérase est l’enzyme responsable de l’ajout de bases nucléotidiques au brin en croissance pendant la réplication de l’ADN. Comme la séquence de nucléotides d’un génome détermine le développement et le fonctionnement d’un organisme particulier, il est vital de synthétiser la réplique exacte du génome existant pendant la réplication de l’ADN. En général, l’ADN polymérase maintient une haute fidélité pendant la réplication de l’ADN, n’incorporant qu’un seul nucléotide mal apparié pour 109 nucléotides ajoutés. Par conséquent, si une erreur d’appariement se produit entre des bases azotées en plus des paires de bases complémentaires standard, l’ADN polymérase ajoute ce nucléotide à la chaîne en croissance, produisant une mutation fréquente. Les erreurs de réplication de l’ADN sont corrigées par deux mécanismes connus sous le nom de relecture et de réparation des mésappariements dirigée vers le brin.
La relecture
La relecture fait référence à un mécanisme initial de correction des paires de bases mal appariées du brin d’ADN en croissance, et elle est effectuée par l’ADN polymérase. L’ADN polymérase effectue la relecture en deux étapes. La première relecture a lieu juste avant l’ajout d’un nouveau nucléotide à la chaîne en croissance. L’affinité des nucléotides corrects pour l’ADN polymérase est plusieurs fois supérieure à celle des nucléotides incorrects. Cependant, l’enzyme doit subir un changement de conformation juste après que le nucléotide entrant se lie à la matrice par des liaisons hydrogène, mais avant la liaison du nucléotide au brin en croissance par l’action de l’ADN polymérase. Les nucléotides mal appariés sont susceptibles de se dissocier de la matrice pendant le changement de conformation de l’ADN polymérase. Cette étape permet donc à l’ADN polymérase de « revérifier » le nucléotide avant de l’ajouter définitivement au brin en croissance. Le mécanisme de relecture de l’ADN polymérase est présenté dans la figure 2.
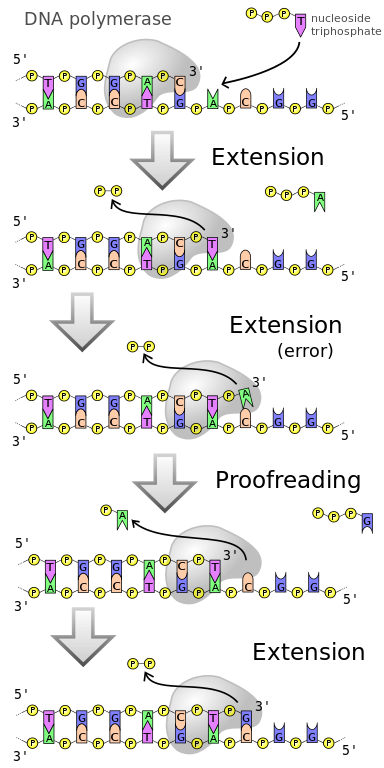
Figure 2 : Relecture
La deuxième étape de relecture est connue sous le nom de relecture exonucléolytique. Elle se produit immédiatement après l’incorporation d’un nucléotide mal apparié au brin en croissance, dans de rares cas. L’ADN polymérase est incapable d’ajouter le deuxième nucléotide à côté du nucléotide mal apparié. Un site catalytique séparé de l’ADN polymérase connu sous le nom d’exonucléase de relecture 3′ à 5′ digère les nucléotides mal appariés de la chaîne en croissance.
Réparation des mésappariements dirigée par le brin
Malgré les mécanismes de relecture, l’ADN polymérase peut encore incorporer des nucléotides incorrects au brin en croissance pendant la réplication de l’ADN. Les erreurs de réplication qui ont échappé à la relecture sont éliminées par la réparation des mésappariements dirigée par le brin. Ce système détecte le potentiel de distorsion de l’hélice d’ADN qui est dû à des paires de bases mal appariées. Cependant, le système de réparation doit identifier la base incorrecte à partir de la base existante avant de remplacer le mismatch. En général, E. coli dépend du système de méthylation de l’ADN pour reconnaître l’ancien brin d’ADN dans la double hélice, car le brin nouvellement synthétisé peut ne pas subir de méthylation de l’ADN rapidement. Chez E.coli, le résidu A du GATC est méthylé. La fidélité de la réplication de l’ADN est augmentée d’un facteur supplémentaire de 102 grâce à l’action du système de réparation des mésappariements dirigé sur les brins. Les voies de réparation des mésappariements de l’ADN chez les eucaryotes, les bactéries et E. coli sont présentées dans la figure 3.
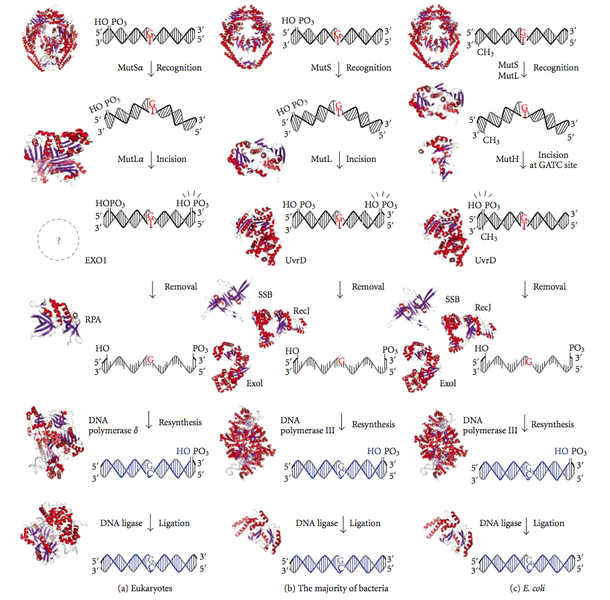
Figure 3 : réparation des mésappariements de l’ADN chez les eucaryotes, les bactéries et E. coli
Dans la réparation des mésappariements dirigée vers le brin, trois protéines complexes se déplacent dans le brin d’ADN nouvellement synthétisé. La première protéine connue sous le nom de MutS détecte et se lie aux distorsions de la double hélice d’ADN. La deuxième protéine, appelée MutL, détecte et se lie à la MutS, attirant la troisième protéine, appelée MutH, qui distingue le brin non méthylé du brin nouvellement synthétisé. Lors de la liaison, la MutH coupe le brin d’ADN non méthylé immédiatement en amont du résidu G de la séquence GATC. Une exonucléase est responsable de la dégradation du brin en aval du mismatch. Cependant, ce système dégrade les régions de moins de 10 nucléotides qui sont facilement re-synthétisées par l’ADN polymérase 1. Les protéines Mut des eucaryotes sont homologues à celle d’E. coli.
Conclusion
Les mutations sont des altérations permanentes de la séquence nucléotidique du génome qui peuvent survenir en raison des erreurs de réplication de l’ADN ou sous l’effet de mutagènes externes. Les erreurs de réplication de l’ADN peuvent être corrigées par deux mécanismes connus sous le nom de relecture et de réparation des mésappariements dirigés vers le brin. La relecture est effectuée par l’ADN polymérase elle-même pendant la synthèse de l’ADN. La réparation des mésappariements dirigée vers le brin est effectuée par les protéines Mut juste après la réplication de l’ADN. Cependant, ces mécanismes de réparation sont impliqués dans le maintien de l’intégrité du génome.
Référence:
1. Alberts, Bruce. « Mécanismes de réplication de l’ADN ». Biologie moléculaire de la cellule. 4e édition, Bibliothèque nationale de médecine des États-Unis, 1er janvier 1970, disponible ici.
2 Brown, Terence A. « Mutation, réparation et recombinaison ». Génomes. 2e édition, Bibliothèque nationale de médecine des États-Unis, 1er janvier 1970, Disponible ici.
Image Courtesy:
1. « Différents types de mutations » Par Jonsta247 – Ce fichier a été dérivé de:Point mutations-en.png (GFDL) via Commons Wikimedia
2. « ADN polymérase » Par I, Madprime (CC BY-SA 3.0) via Commons Wikimedia
3. « DNA mismatch repair » Par Kenji Fukui – (CC BY 3.0) via Commons Wikimedia
![]()
.